Де у людини найбільше тактильних рецепторів. Класифікація та структура рецепторних утворень шкірного аналізатора. Фізіологія мозочка, його вплив на моторні та вегетативні функції організму
У соматосенсорну систему включають систему шкірної чутливості та чутливу систему скелетно-м'язового апарату, головна роль якої належить пропріорецепції.
1) Отримання інформації від рецепторів.
2) Переробка інформації про різні подразники.
Мехонарецептори
Ноцирецептори
Терморецептори
Пропріорецетори
Швидко адаптуються: рецептори волосяних фолікул, Тельця Пачіні, Тельця Мейсснера, Колбочки Краузе, Вільні н. закінчення типу А?.
Повільно адаптуються: Диски Меркеля, Тельця Руфіні, вільні зв. закінчення типу С.
Рецептори волосяних фолікулів
Розташування: у внутрішньому шарі шкіри, оточують волосяну цибулину
Адаптація: швидка. Розряд припиняється через 50-500 мс після включення стимулу
Рецепція: на рух, посмикування волосків, але не на ступінь їх зміщення
Іннервація: одне нервове волокно може обслуговувати кілька сотень фолікулів, а кожен фолікул може іннервуватися безліччю рецепторів
Тільця Пачіні (пластинчасте тільце, Фатера-Пачіні тільце)
Будова: має будову цибулини або матрьошки. Укладено у слизові оболонки із сполучної тканини. Усередині розташоване еліптичне нервове віконце.
Розмір: 0,5 – 0,7 мм у поперечнику та близько 1-2 мм у довжину
Розташування: як у волосистій так і в гладкій шкірі, глибокі слові шкіри (у жировій тканині підшкірних шарів, глибше інших розчинів), мало в губах, подушечках пальців
Рецепція: сильні та різкі зміни тиску на шкіру. Чи не реагують на постійний тиск. Вібрація: реагують на вібрацію від 70 Гц до 1000 Гц. Однак найбільша чутливість на частоті вібрації 200-400 Гц, у цьому випадку здатні реагувати на деформацію шкіри лише на 1 мкм.
Тільця Пачіні не відключаються при місцевій анестезії
Тельця Мейснера
Розташованіу поверхневих шарах гладкої шкіри (сосочковий шар дерми) та на слизових оболонках. Найбільше їх на губах, долонях, пальцях, підошвах
Є аналогом рецепторів волосяних фолікулів для гладкої шкіри.
Будова: капсула із сполучної тканини овальної форми (довжина 40-180 мкм, ширина 30-60 мкм)
Нервові закінчення утворюють всередину капсули спіраль, гілки якої ізольовані один від одного оболонками шванновських клітин.
Капсула прикріплена до вищележачих шарів епітелію колагеновими волоками (що підвищує механічний зв'язок між нею та поверхнею шкіри)
Рецепція: реагують на дотик або тиск
Швидко адаптуються. Розряд припиняється через 50-500 мс після включення стимулу
Реагують на низькочастотну вібрацію 10-200 Гц максимум на частоті 30 Гц.
Мають малі рецептивні поля
Колбочки Краузе (термінальні колби Краузе, цибулини Краузе)
Розташування: епідерміс гладкої шкіри та слизові оболонки. Є тільки у ссавців, які не належать до приматів (у людей немає)
Будова:схожі з тільцями Мейснера. Пластинчаста капсула, всередині якої спіралеподібне або стрижнеподібне нервове закінчення
Рецепція: довгий час вважалося, що це розчини холоду, але це не так. Колби Крауз реагують на низькочастотну вібрацію 10-100 Гц.
Тактильні рецептори, що повільно адаптуються.
1) Диски Меркеля
2) Тельця Руффіні
3) Вільні нервові закінчення (тип С)
Ілюзія сенсорного розмаїття
Диски Меркеля
Розташування: на ділянках гладкої шкіри розташовуються невеликими групами в нижніх шарах епідермісу, звідки прямують у сосочки дерми. На волосистих ділянках розташовуються у спеціальних тактильних дисках (тільця Пінкуса-Ігго) – невеликих піднесення шкіри.
Будова: капсули з великими неправильною формою ядрами та мікроворсинками
Іннервація: на три тактильні диски може припадати одне нервове волокно, а всередині тактильних дисків всі диски Меркеля (30-50 шт) обслуговуються першою нервовою гілкою.
Рецепція: реагують на дотик чи тиск. Стимул - прогинання епідермісу при дії механ стимулу. Повільно адапт.р-ри. Продовжують генерувати потенціали навіть коли тиск підтримується довгий час. Мають малі рецептивні поля.
Тільця Руффіні (Циліндри Руффіні, закінчення Руффіні)
Розташування: нижній шар дерми та слизової оболонки.
Рецепція: вважалося, що вони реагують на тепло, але це не так Реагують на тривале усунення шкіри, тиск.
Повільно адаптований продовжують генерувати птенціали, навіть коли тиск підтримується довгий час.
Мають великі рецепторні поля.
Вільні нервові закінчення
Розташування: в епідермісі та дермі, найпоширеніші розчини. Виявлено практично у всіх ділянках шкіри.
Будова: немає спеціалізованих детекторних кл-ок. від закінчень відходять волокна типу А дельта (мієлінізовані) або типу С(немієлінізовані).
Рецепція: збуджуються при дуже слабкому, пороговому подразненні. Реагують тільки на 1 градацію стимулу (є-ні). Можуть детектувати слабкі механ стимули, рух по шкірі (комаха повзуча)
Адаптація: волокна типу А дельта ...?
Передній спиноталамічний шлях – див.
· Перший нейрон – аксони у складі задніх корінців входять у задній ріг СПМ, тіло у СПМ ганглії, дендрит закінчується механорецепторами шкіри
· Другий нейрон - аксони переходять на інший бік СПМ і утворюють передній спинотаамічний тракт, тіло і дендрити в кл-ках драглистого реч-ва (задній ріг СПМ)
· Третій нейрон – аксони: частина у постцентральну звивину, частина у верхню тім'яну часточку, тіло та дендрити у задніх вентролатеральних ядрах таламуса
Пропріорецепція.
Пропріоцепція є сприйняття пози і руху нашого власного тіла. Поза визначається кутом розташування кісток у кожному суглобі, що встановлюється або пасивно (зовнішніми силами), або активно (м'язовим скороченням). З їхньою роботою поєднуються сигнали від вестибулярного органу, що дозволяє визначати положення тіла у полі земного тяжіння. Пропріорецептори беруть участь також у нашій свідомій та несвідомій руховій активності. Аферентні та еферентні системи у поєднанні створюють усвідомлювані пропріоцептивні відчуття. Якщо відчуття, наприклад, рух у суглобі зберігається після того, як один із компонентів системи усунений, з цього не обов'язково випливає, що він у нормі не бере участі у формуванні даного відчуття. Це відповідає принципу надмірності нервової системи. Аферентна інформація може модулюватися в синапсах низхідним гальмуванням.
У синапсах, через які активність аферентів передається центральному соматосенсорному нейрону, воно може змінювати величину рецептивного поля цього нейрона, якщо аференти, що йдуть від периферичної частини рецептивного поля, гальмуються.
Види пропріорецепторів
Ссавці:
1) М'язові веретени
М'язові волокна ссавців
1) Екстрафузальні. Виконують усю роботу скорочення м'язів
2) Інтрафузальні. Вони відсутні актин і міозин. Вони призначені для детектування натягу за допомогою розчинів, званих м'язові веретени
· Статичні. Реагують за постійного натягу м'яза. Детектують силу скорочення
· Динамічні. Реагують на вкл-викл розтягування м'язів. Детектують швидкість скорочення
2) Сухожильні органи Гольджі
У сухожиллях- Частина м'яза, представляє собою сполучнотканинну формацію, за допомогою якої м'язи прикріплюється до кістки.
СОГ – грознеподібні чутливі закінчення (2-3 мм завдовжки і 1-1,5 мм завширшки). Порушуються при скороченні м'язів через натяг сухожилля.
3) Рецептори суглобів
· У суглобових сумках: закінчення на кшталт тілець Руффіні. Повільно адаптуються. Кожне має свій «кут порушення»
· У суглобових зв'язках: закінчення на кшталт тілець Гольджі та тілець Пачіні. Активуються під час руху суглоба до крайніх положень чи його обертання виходить межі норми.
Нервові шляхи
1) Корковий пропріоцептивний шлях- точно локалізовані усвідомлені пропріорецептивні відчуття
· Шлях Бурдаха
· Шлях Голля
Його поразки:
1. Втрата почуття становища та локомоції. При закритих очах пацієнт не може визначити положення своїх кінцівок
2. Астереогноз. При закритих очах хворий може дізнатися та описати предмет на дотик.
2) Шляхи мозочкового спрямування- Неусвідомлена координація рухів
· Шлях Флексінгу
· Шлях Говерсу
Поразки цих шляхів: розлад координації рухів. Стає неможливо без зорового контролю виконати будь-який навіть найпростіший рух без грубих помилок. Наприклад, торкнутися кінчика носа.
Схема тіла
Схема тіла- Неусвідомлювані уявлення про становище власного тіла та його частин у просторі, про його межі та динамічних хар-ках.
Властивості схеми тіла (за Хаггардом і Волпертом)
1) Просторове кодування
3-мірні просторові координати тіла та об'єктів навколо. Уявлення про межі тіла може не відповідати його реальним кордонам (теніс – відтворення продовження тіла як кінець ракетки).
2) Модульність
Схема тіла не представлена в якійсь єдиній області мозку. Різні частини тіла – у різних областях кори.
3) Адаптивність
Уявлення про схему свого тіла розвиваються в ході життя.
Соматосенсорна пластичність
4) Оновлюваність під час руху
Після виконання рухів схема тіла змінюється згідно з новим положенням тіла
5) Інтерперсональність
Св із дзеркальними нейронами.
6) Надмодальність
Олівер Сакс. «Людина, яка випала з ліжка» Схема тіла не пов'язана з опр сенсорною модальністю. Вона включає пропріорецепцію, зір, тактильну інформацію та ін. Сенсорна інформація перекодується в абстрактну, надмодальну форму.
7) Когерентність
p align="justify"> При формуванні схеми тіла інтегрується інформація від різних органів чуття.
Подібна інформація.
Соматосенсорна системапоєднує нервові механізми, що збирають сенсорну інформацію від тіла. Соматичні відчуття протистоять специфічним почуттям, до яких відносять зір, слух, запах, смак та рівновагу.
Можна виділити три фізіологічні типу соматичних відчутті: (1) механорецептивні, що включають тактильні відчуття та почуття становища; вони стимулюються механічними усуненнями деяких тканин тіла; (2) терморецептивні, що виникають під дією тепла та холоду; (3) больові, виникнення яких пов'язане з дією будь-якого фактора, що ушкоджує тканини.
Інші класифікації соматичних відчуттів. Існують також інші класифікації соматичних відчуттів: (1) екстероцептивні відчуття, що виникають при подразненні рецепторів на поверхні тіла; (2) пропріоцептивні відчуття, що мають відношення до фізичного стану тіла, включаючи положення тіла, стан м'язів та сухожиль, ступінь тиску на підошви стоп і навіть відчуття рівноваги (яке часто розглядають як специфічне, а не соматичне відчуття).
Вісцеральні відчуттяпов'язані зі станом внутрішніх органів. Глибокі відчуття походять від глибоких тканин, наприклад фасцій, м'язів та кісток. До цих відчуттів відносять головним чином «глибокий» тиск, біль та вібрацію.
Взаємини між відчуттямидотику, тиску та вібрації. Хоча дотик, тиск і вібрацію часто вважають окремими відчуттями, всі вони сприймаються одними й тими самими типами рецепторів. Існують три основні відмінності між ними: (1) відчуття дотику зазвичай виникає при стимуляції тактильних рецепторів у самій шкірі або тканинах, розташованих відразу під нею;
(2) відчуття тиску викликає деформація глибших тканин;
(3) вібрація відчувається при дії швидко повторюваних сигналів в результаті збудження деяких рецепторів, що сприймають дотик і тиск.
Тактильні рецептори. Відомі принаймні шість абсолютно різних типів тактильних рецепторів, але набагато більше їх різновидів. Деякі з цих рецепторів показані малюнку; їх функціональні особливості представлені далі.
Перший тип рецепторів- Вільні нервові закінчення. Вони повсюдно поширені в шкірі та багатьох інших тканинах і реагують на дотик і тиск. Наприклад, навіть легкий контакт з роговою оболонкою ока, в якій немає інших рецепторів, крім вільних нервових закінчень, проте викликає відчуття дотику і тиску.
Другий тип рецепторів- Високочутливе дотик тільце Мейсснера. Воно являє собою подовжене інкапсульоване закінчення чутливого мієлінового нерва типу А. Усередині капсули нервове закінчення розгалужується на безліч волоконець. Тельця Мейсснера локалізуються в позбавлених волосяного покриву ділянках шкіри, але особливо багато в кінчиках пальців рук, губах та інших областях, де в людини високо розвинена здатність до просторового розрізнення тактильного подразнення. Тельця Мейсснера адаптуються протягом частки секунди від початку їхньої стимуляції, тому вони особливо чутливі до руху об'єктів по поверхні шкіри, а також до вібрації низької частоти.
Третій тип – диски Меркеля. Їх відносять до тактильних рецепторів із розширеним кінчиком. Значна кількість цих рецепторів зустрічається в кінчиках пальців рук та інших позбавлених волосяного покриву ділянках шкіри, що містять велику кількість тілець Мейсснера. Волосисті ділянки шкіри також містять помірну кількість рецепторів з розширеним кінчиком, хоча тут майже немає тілець Мейсснера. Диски Меркеля відрізняються від тілець Мейсснера характером адаптації: спочатку в них виникає сильний сигнал, що частково адаптується, потім в рецепторі тривалий час зберігається більш слабкий, дуже повільно адаптується сигнал. Отже, ці рецептори відповідальні за передачу стабільних сигналів, що дозволяють відчувати безперервний контакт об'єктів зі шкірою.
Диски Меркелячасто групуються разом, формуючи рецепторний орган, званий куполом Ігго, який упирається в шкіру з нижнього боку епітелію. Епітелій у цій точці видається назовні, формуючи купол, що містить надзвичайно чутливий рецептор. Зверніть увагу, що вся група дисків Меркеля іннервується одним товстим нервовим мієлінізованим волокном (тип А). Ці рецептори поряд з тільцями Мейсснера відіграють важливу роль у локалізації тактильних відчуттів на поверхні тіла та у визначенні поверхневої структури об'єкта, що відчувається.
Четвертий тип включає рецептори, відповідальні за відчуття, що виникають під час руху будь-якого волосся на тілі. При цьому стимулюється нервове волокно, що обвиває основу волосся, яке називають волосяним рецептором. Таким чином, кожне волосся та нервове волокно в його основі є тактильним рецептором. Цей рецептор швидко адаптується і подібно до тільців Мейсснера реагує головним чином на (а) рух об'єктів по поверхні тіла або (б) первинний контакт подразника з тілом.
До п'ятого типуможна віднести рецептори Руффіні – інкапсульовані, сильно розгалужені закінчення. Їх багато у глибоких шарах шкіри, а також у ще глибших внутрішніх тканинах. Ці рецептори адаптуються дуже повільно і, отже, важливі передачі інформації про безперервну деформацію тканин, що виникає, наприклад, при інтенсивному тривалому дотику і тиску. Рецептори Руффіні виявлені також у суглобових сумках та допомагають визначати ступінь обертання суглоба.
Шостий тип - тільця Пачіні. Вони лежать як безпосередньо під шкірою, так і глибоко у фасціальних тканинах тіла. Тільця стимулюються лише швидким локальним стиском тканин, оскільки адаптуються протягом кількох сотих часток секунди, тому дані рецептори особливо важливі для сприйняття вібрації тканин або інших швидких змін їх механічного стану.
У шкірі людини є тактильні (дотикові), температурні та болючі рецептори. Різні види рецепторів відрізняються за своєю будовою та розподілені у шкірі у вигляді своєрідної мозаїки.
Тактильні рецептори сприймають механічні подразнення, що супроводжуються почуттям дотику та тиску. Вони мають форму подовжених цибулин, до яких підходять нервові закінчення. До тактильних рецепторів відносяться:дотичні тільця (тільця Мейснера) мають вигляд одного звивистого нервового закінчення, одягненого в капсулу; пластинчасті тільця (тільця Пачіні), що складаються з нервового закінчення, оточеного сполучнотканинними пластинками; дотичні диски Меркеля, розміщених біля волосяних цибулин, в епідермісі, а також у судин і в глибоких шарах шкіри поверхні кисті, в області долонь, а також на кінчиках пальців, губ, сухожиль, очеревині та брижі кишечника і т.д.. У середньому на 1 см2 шкіри припадає 25 тактильних рецепторів.
Більше рецепторів у шкірі долонь, на кінцях пальців, на губах та кінчику язика; найменше — у шкірі спини та живота. Поріг подразнення найбільш чутливих ділянок становить 50 мг, а в найменш чутливих - до 10 г. За функціональними особливостями тактильні рецептори поділяють на фазні та статичні Фазні тактильні рецептори збуджуються при динамічному подразненні, мають високу чутливість, короткий латентний період і швидко адаптуються. Статичні тактильні рецептори збуджуються в основному від статичних подразнень, вони менш чутливі, але мають триваліший латентний період і повільніше адаптуються.
Порушення, яке виникає в тактильних рецепторах при контактах шкіри з предметами, надходить до мозкового центру тактильного аналізатора, що локалізується в ділянці І сомато-сенсорної зони кори головного мозку (задньої центральної закрутки кори великих півкуль), де трансформується у відчуття дотику або тиску. Диференціювання цього відчуття залежить від адаптаційних здібностей тактильних рецепторів шкіри:як зазначалося вище, легко адаптуються фазні тактильні рецептори і вони реагують лише зміну інтенсивності стимулу і дають короткочасне відчуття дотику, навіть якщо стимул тиску діє тривало. Статичні тактильні рецептори, що адаптуються повільно, збуджуються тільки при тривалому впливі механічного подразника, чим і забезпечують відчуття тиску. За механізмом дотику можуть сприйматися і подразнення у вигляді вібрації. Завдяки тактильній чутливості людина відчуває форму, величину та характер поверхні навколишніх предметів. Для дотику характерно також просторове відчуття, яке полягає у здатності розрізняти і сприймати як окремі дві одночасно роздратовані точки тіла.
До терморецепторів, або температурних рецепторів належать два види нервових закінчень. Одні (колбочки Краузе) сприймають переважно холодові подразнення, а другі (тільця Руффіні) — теплові подразнення. Розміщені терморецептори в шкірі, а також у слизовій оболонці носа, рота, гортані, стравоходу, шлунка та кишечника. За будовою терморецептори є клубочками тонких нервових закінчень, які містяться в сполучнотканинних капсулах. подразнює терморецептори шкіри та викликає в пробковій частині аналізатора відчуття тепла чи холоду. Внаслідок цього рефлекторно змінюється просвіт кровоносних судин шкіри, завдяки чому змінюється її кровопостачання та температура.
Холодових рецепторів в організмі близько 250 тис., теплових – до 30 тис. – холодові рецептори розташовані на глибині 0,17 мм, а теплові – 0,3 мм від поверхні шкіри. Завдяки цьому теплові рецептори збуджуються відносно повільно, тоді як холодові реагують дуже швидко, як на подразнення з температурою нижче 18-20°С та на подразнення температурою вище 40-45°С (наприклад, ефект «гусячої шкіри» при зануренні тіла у гарячу воду) ). Терморецептори постійно інформують організм про стан та зміну температури навколишнього середовища та є найважливішою ланкою у підтримці температури (термостазу) тіла. Діти температурне почуття проявляється вже з перших днів після народження.
Біль є специфічним почуттям, якісно відмінним від будь-якого іншого почуття. Воно виникає тоді, коли на ту чи іншу частину організму діє подразник, що має руйнівний характер. У цьому виникає ціла черга захисних реакцій, вкладених у збереження частин тіла чи цілого організму.
Больові подразнення сприймаються больовими рецепторами або вільними нервовими закінченнями. Больові рецептори розташовані у шкірі, а й у м'язах, кістках, внутрішніх органах. На поверхні 1 см2 буває близько 100 болючих точок, а на всій поверхні шкіри їх близько мільйона. На шкірі майже немає ділянки, де не було б больових рецепторів, але розташовані вони нерівномірно: більше в пахвовій та пахвинній ділянках і найменше – на підошвах, долонях, вушних раковинах. Порушення, що виникають у больових рецепторах внаслідок дії подразника, передаються по доцентрових нервах у вищі кіркові та підкіркові (у таламусі та гіпоталамусі) центри болю, де і формуються відчуття болю. Сила больових відчуттів залежить від стану нервової системи. Больові рецептори реагують на значні коливання температури, тиску, концентрацію простагландинів, виділених пошкодженими клітинами тіла. Попадання інформації про локалізації та інтенсивність болю до центрів головного мозку стимулює виділення в кров ендорфінів, які є блокаторами болю.
При болючих подразненнях рефлекторно порушується нормальна діяльність організму і особливо:посилюється виділення адреналіну в кров, підвищується концентрація цукру в крові, порушується ритм серцевих скорочень, прискорюється згортання крові, підвищується кров'яний тиск, затримується дихання та ін.
Ще одним видом шкіряної чутливості є сприйняття лоскотання, що забезпечують вільно розташовані в поверхневих шарах шкіри нервові закінчення. Для цього виду рецепторів властиві специфічні реакцію стимули різної інтенсивності. З активацією цієї групи рецепторів раз і пов'язують відчуття лоскоту, що й дало назву самим рецепторам - рецептори лоскотання.
Внаслідок дії термічних факторів, хімічних речовин, електричного струму або іонізуючого випромінювання можуть виникнути пошкодження тканин організму і, насамперед, шкіри, які називаються опіками. По глибині пошкодження тканин розрізняють чотири ступені опіків. Опіки І ступеня характеризуються місцевим (ерітелію), незначною припухлістю та підвищенням місцевої температури, триває 2-5 днів і звично проходить безвісти. Опіки II ступеня також викликають місцеве почервоніння та припухлість шкіри, а крім того, ще й характеризуються появою бульбашок, наповнених рідиною (лімфою) жовтого кольору. Такі опіки супроводжуються болем та лихоманкою.
Опіки III-А ступеня супроводжуються Ш-Б ступеня некрозом всіх шарів шкіри, а IV ступеня - некрозом шкіри та глибоких тканин. Невідкладна допомога при опіках полягає в негайному видаленні та нейтралізації фактора, що викликав. При опіках хімічною речовиною уражену шкіру та слизові оболонки слід негайно промити великою кількістю холодної проточної води (протягом не менше 15 хв). У разі опіку шкіри сірчаною кислотою або негашеним вапном промивати уражене місце водою не можна, оскільки вона лише посилить їхню дію. Для цього використовують олію або тваринну олію. При тяжких ураженнях хворих госпіталізують.
Тельця Мейсснера, розташовані в поверхневих шарах власне шкіри (дерми) губ та власне слизової оболонки рота, реагують на дотик. При посиленні механічного подразнення порушуються диски Меркеля, які локалізуються у глибоких шарах шкірного епідермісу та слизового епітелію Відчуття тиску та вібрації виникають при подразненні тілець Пачіні, розташованих у підшкірній клітковині та підслизовому шарі. У зв'язку з глибоким заляганням тілець Пачіні, місцева аплікаційна анестезія поверхневих шарів слизової оболонки та шкіри не усуває відчуття тиску та вібрації, про що необхідно попередити пацієнта перед операцією в цих умовах.
Від більшості тактильних механорецепторів ротового відділу сенсорні сигнали надходять до ЦНС мієлінізованим нервовим волокнам Аb зі швидкістю 30-70 м/с. Центральний відділ тактильної сенсорної системи розташовується в задній звивині центральної кори великих півкуль.
Тактильні відчуття можна викликати роздратуванням тільки певних ділянок шкіри та слизових, які називаються чутливими тактильними точками . Просторовий поріг тактильної чутливості обернено пропорційний кількості рецепторів на одиницю площі і прямо пропорційний відстані між рецепторами. Просторовий поріг тактильних відчуттів на кінчиках пальців, язика та губ значно нижче (1-3 мм), ніж на інших ділянках тіла (50-100 мм). Це зумовлено відмінністю густини рецепторів на одиницю поверхні.
Найбільш щільно тактильні рецептори розташовані на кінчику язика, слизовій оболонці та червоній облямівці губ, що необхідно для апробації їжі на їстівність. Найбільш чутлива до механічних подразнень верхня губа. Відносно високим рівнем тактильної чутливості відрізняється слизова оболонка твердого піднебіння, що забезпечує формування харчової грудки в процесі жування. Найменшою тактильною чутливістю має слизова оболонка вестибулярної поверхні ясен. При цьому в області ясенних сосочків відзначається спадний градієнт чутливості від різців до молярів.
Метод дослідження абсолютних або просторових порогів тактильної чутливості естезіометрією . Вивчення тактильного сприйняття слизової оболонкою ротової порожнини дозволяє прогнозувати індивідуальні особливості адаптації до знімних зубних протезів у хворих з частковою або повною адентією. Протез є стороннім тілом, що подразнює тактильні рецептори, що веде до рефлекторної гіперсалівації, виникнення блювотного рефлексу, порушення координації жування, ковтання та мови. Однак більшість тактильних рецепторів відноситься до швидкоадаптованих. У зв'язку з цим, а також внаслідок відсутності тактильних рецепторів, що не адаптуються, істотних проблем зі звиканням до зубних протезів, як правило, не виникає. При цьому поряд із пристосуванням рецепторного апарату відбувається адаптація провідникового та центрального відділів аналізатора. Це є результатом високої пластичності нервових центрів, що забезпечують швидке пристосування функцій жування, ковтання та промови до нових умов. Зубний протез перестає відчуватися як стороннє тіло, спостерігається відновлення ефективності жування, згасає блювотний рефлекс, нормалізуються салівація, ковтання та мова.
Температурна рецепція у ротовому відділізабезпечує сприйняття термічних подразників - тепла та холоду. Терморецептори, що сприймають холод, гістологічно представлені колбами Краузе, розташованими в епідермісі червоної облямівки губ та епітелії слизової оболонки рота. Теплові рецептори – тільця Руффіні, локалізуються глибше – у власне дермальному шарі губ та у власне слизовій оболонці рота. Від рецепторів холоду відходять тонкі мієлінізовані волокна типу Аd зі швидкістю проведення збудження 5-15 м/с, а від рецепторів тепла безмієлінові волокна типу С (0,5-3 м/с). Центральний відділ температурної сенсорної системи розташовується у задній центральній звивині кори великих півкуль.
Як правило, теплові та холодові рецептори збуджуються відповідними за якістю стимулами. Однак за певних умов холодові рецептори можуть сприймати теплові подразники при температурі понад 45 0 С (наприклад, при зануренні у гарячу ванну). Залежно від вихідних умов, та сама температура може викликати і відчуття тепла і відчуття холоду.
Переважання у шкірі та слизових терморецепторів, що реагують на холодові стимули (10:1), та глибоке залягання теплових рецепторів, зумовлюють більш високу чутливість до холоду. При цьому холодова чутливість знижується від передніх відділів рота до задніх, а теплова, навпаки, підвищується. Найбільшою чутливістю до температурних подразнень відрізняються кінчик язика та червона облямівка губ, що забезпечує апробацію придатності споживаної їжі. Малочутлива до холоду та тепла слизова оболонка щік. Повністю відсутнє сприйняття тепла у центрі твердого піднебіння, а центральна частина задньої поверхні язика не сприймає ні теплові ні холодові дії.
Здатність до сприйняття температури мають рецептори дентину і пульпи зубів. Поріг холодової чутливості для різців в середньому становить 20 0 С, а для іклів, премолярів та молярів – 11-13 0 С. Порогом теплової чутливості для різців є температура близько 52 0 С, для решти зубів – 60-70 0 С.
Дослідження температурної чутливості шляхом визначення теплових чи холодових порогів називають термоестезіометрією . Для дослідження температурної чутливості зубів їх зрошують гарячою або частіше холодною водою або використовують ватний тампон, змочений в ефірі, який, випаровуючись, охолоджує зуб. Якщо температурні подразники викликають адекватні відчуття тепла чи холоду, це свідчить про нормальний стан тканин зуба. При карієсі холодове подразнення спричиняє біль. При пульпіті біль викликають теплові стимули, а холодові навпаки зменшують її. Депульпований зуб не реагує на холод, ні тепло.
Тактильна та температурна чутливістьротового відділудоповнюється м'язово-суглобової рецепціїщо забезпечує почуття просторового становища нижньої щелепи щодо верхньої, відчуття її руху, сприйняття скорочувального зусилля м'язів. Цей вид чутливості забезпечується пропріорецепторами, які локалізуються в інтрафузальних м'язових волокнах, скронево-нижньощелепних суглобах, у зв'язковому апараті жувальних та мімічних м'язів. Сенсорні сигнали від пропріорецепторів надходять до ЦНС переважно по товстим мієлінізованим нервовим волокнам типу Аa зі швидкістю 70-120 м/с. Центральний відділ пропріоцептивної сенсорної системи розташовується у задній центральній звивині кори великих півкуль.
Найважливішою сенсорною функцією ротового відділу є больова рецепціяяка забезпечує сприйняття стимулів, здатних призвести до пошкодження або руйнівних тканин організму. На відміну від інших видів сенсорних модальностей больова рецепція немає адекватного подразника. Практично будь-який надсильний стимул може спричинити відчуття болю.
Біль- це універсальне неприємне сенсорне відчуття і емоційне переживання, пов'язане з загрозою руйнування або пошкодженням тканини, що вже відбулося.
Відповідно до біологічної значущості розрізняють два види болю: фізіологічнуі патологічну. Основні завдання фізіологічного болю:
1) інформування організму про будь-які форми загрози його існуванню чи цілісності,
2) участь в організації адаптивної поведінки, спрямованої на запобігання поширенню та ліквідацію пошкодження або усунення її загрози.
Біль забезпечує мобілізацію більшості систем організму для захисту від пошкодження тканин та супроводжується розгортанням оборонної поведінки. Залежно від ситуації відчуття болю і поведінкові та рефлекторні реакції, що супроводжують її, можуть свідомо пригнічуватися. Проте, гуморальні, і навіть вегетативні зрушення зберігаються у разі, що є неминучою ознакою ушкодження тканин. Тому при усуненні больових синдромів доцільно використовувати лікарські препарати, здатні стабілізувати фізіологічні функції організму.
Після організації захисної поведінки біль втрачає свої адаптивні функції та набувають значення самостійного патогенетичного фактора. Для багатьох захворювань біль – один із перших, а іноді й єдиний прояв патології та головний діагностичний індикатор.
За місцем застосування ушкоджуючого фактора виділяють два роди болі: соматичнуі вісцеральну. Соматичний біль пов'язаний з екстремальними зовнішніми впливами, а вісцеральний обумовлений внутрішніми патологічними процесами.
Соматичний більподіляється на два типи: первиннуі вторинну. Первинна (епікритична )біль проявляється відразу після пошкодження, швидко усвідомлюється, легко детермінується за якістю та локалізації, зникає після припинення шкідливої стимуляції, супроводжується адаптацією. Вторинна (протопатична )біль проявляється через 0,5-1 з після первинного відчуття, повільно усвідомлюється, погано детермінується за якістю та локалізації, зберігається тривалий час після припинення стимуляції, не супроводжується адаптацією.
В даний час є три основні теорії механізмів сприйняття болю:
1) теорія інтенсивності,
2) теорія специфічності,
3) теорія розподілу імпульсів.
Відповідно до теорії інтенсивності надсильна стимуляція рецепторів, незалежно від їх модальності, викликає високоамплітудні РП та високочастотну розрядну діяльність сенсорних нейронів, що трасформується ЦНС у відчуття болю (амплітудно-частотне кодування).
Відповідно до теорії розподілу імпульсів ушкоджуючі стимули викликають особливий порядок слідування (паттерн) аферентних імпульсів, який відрізняється від розрядної діяльності, викликаної індиферентними для організму факторами (інтервально-імпульсне кодування). При цьому ЦНС перетворює аферентний потік, що надходить у відчуття болю.
На противагу цьому теорія специфічності передбачає (за аналогією з іншими сенсорними системами) існування спеціальних рецепторів та аферентів, що реагують збудженням тільки на такі за інтенсивністю стимули, які можуть пошкодити тканину (двійкове та просторове кодування).
Таким чином, подразник може викликати відчуття болю тільки в тому випадку, якщо під його впливом формується особлива, алгогенна сигналізація- потік аферентних збуджень, в якому за амплітудно-частотно-просторовим принципом закодована інформація про загрозу руйнування або пошкодження тканин організму, що вже відбулося.
Сенсорна система, що забезпечує сприйняття шкідливих подразників, називається ноцицептивний . Рецептори цієї системи ноцицептори, Поділяються на чотири види:
1) механочутливі, які збуджуються в результаті механічного зміщення рецепторної мембрани,
2) хемочутливі, що реагують на хімічні речовини, що виділяються пошкодженими клітинами (ацетилхолін, гістамін, серотонін, простагландини),
3) термочутливі, які активуються під впливом термічних стимулів, що виходять за межі фізіологічного діапазону,
4) полімодальні, що реагують як на хімічні речовини, так і на інтенсивні механічні та термічні стимули.
Ноцицептори відносяться до неадаптованих, високопорогових рецепторів. У шкірі обличчя та слизової оболонки рота, а також періодонті, пульпі та дентині зубів вони переважно представлені вільними нервовими закінченнями.
Вираженою больовою чутливістю відрізняється слизова оболонка вестибулярної поверхні нижньої щелепи в ділянці бічних різців. Найменшою больовою чутливістю характеризується язична поверхня слизової оболонки ясен. На внутрішній поверхні щоки в області верхніх молярів є вузька ділянка слизової оболонки, абсолютно позбавлена больової чутливості.
Винятково сильне больове відчуття виникає навіть при легкому дотику до пульпи зуба, що обумовлено високою щільністю високочутливих нервових закінчень та волокон, які проникають до дентину аж до емалево-дентинної межі. На 1 см 2 дентину припадає 15000-30000 больових рецепторів, на межі емалі і дентину кількість ноцицепторів доходить до 75000, тоді як у шкірі їх число не перевищує 200. Все це є причиною особливої жорстокості болю, що виникає під впливом температурних, хімічних при пошкодженні та руйнуванні тканин зубів, у тому числі і при їх лікуванні.
Сенсорні сигнали від ноцицепторів ротового відділу надходять у ЦНС по мієлінізованим нервовим волокнам типів Аb і Аd, а також безмієліновим волокнам групи С, більшість яких проходить у складі другої і третьої гілки трійчастого нерва. Інформація від ноцицепторів про неблагополуччя тканин ротового відділу надходить у задню центральну звивинута до медіальних відділів орбітальної коривеликих півкуль.
Тісний взаємозв'язок між різними ядрами трійчастого нерва та їх взаємодія з ядрами ретикулярної формації обумовлює широку іррадіацію збудження, що ускладнює локалізацію зубного болю та її відображення (проеціювання) у досить віддалені ділянки обличчя, голови та шиї.
Іноді після операції видалення ураженого зуба зберігається відчуття болю, який називається фантомний . Фантомні болі обумовлені тим, що ноцицептивна аферентація від ураженого зуба, що передує видаленню, викликає нейрогенну (центральну ) сенситизацію - збільшення чутливості, пов'язане з підвищенням збудливості у провідниковому та центральному відділах ноцицептивної системи. Додаткове подразнення під час операції викликає появу стійких патологічно посилених вогнищ циркуляції збудження в ЦНС, яке сприймається клітинами кори мозку як тривалий, часто безперервний біль. Лікувальні заходи місцевого характеру не призводять до зменшення або припинення таких болів, оскільки їхнє джерело лежить у структурах ЦНС, на які слід впливати, активуючи антиноцицептивну систему мозку.
Основні функції ендогенної антиноцицептіаної системи – обмеження рівня больового збудження, а також регуляція та підтримання порога больової чутливості. Це забезпечується за рахунок механізмів пресинаптичного та постсинаптичного гальмування ноцицептивних нейронів на всіх рівнях ЦНС. У реалізації впливу антиноцицептивної системи беруть участь опіатні, адренергічні, дофамінергічні та серотонінергічні структури мозку. Провідне значення при цьому має вироблення опіатних морфіноподібних сполук – ендорфінів, енкефалінів та динарфінів.
Больовий поріг є результатом взаємодії ноцицептивної та антиноцицептивної системи, яка перебуває у стані постійної тонічної активності. Усунення постійного гальмівного впливу антиноцицептивної системи може призвести до стану гіпералгіїабо навіть виникнення мимовільних больових відчуттів. Підвищення тонічної активності антиноцицептивної системи призводить до розвитку вроджених аналгій- Нечутливість до болю.
Страх, пригнічуючи активність антиноцицептивної системи, різко посилює реакцію на біль, знижують поріг больової чутливості, а стан типу агресії-люті, навпаки, збільшує його. Переоцінка інтенсивності болю може бути пов'язана з підготовкою та очікуванням медичних маніпуляцій. Проте больова чутливість знижується, коли людина заздалегідь попереджений характер майбутнього впливу. Роз'яснення або відволікаючі розмови перед операцією суттєво послаблює больові відчуття та знижують потребу у знеболювальних засобах.
Специфічною особливістю сенсорної функції ротового відділу є смакова чутливість.
Смак- відчуття, що виникає в результаті сприйняття чотирьох елементарних смакових якостей хімічних речовин, розчинених у ротовій рідині. солодкого, гіркого, кислогоі солоного.
Сенсорна система, яка здійснює контактне сприйняття та оцінку смакових властивостей хімічних речовин, що діють на орган смаку, називається смаковим аналізатором .
Орган смаколюдинипредставлений смаковими ниркамиякі локалізуються, переважно, в сосочках мови: грибоподібних, листоподібнихі жолобоподібних. Грибоподібні сосочки розташовуються, головним чином, на слизовій оболонці кінчика язика, листоподібні сосочки - вздовж бічної поверхні задніх відділів язика, а жолобоподібні - поперек спинки, біля кореня язика. Окремі смакові бруньки є на м'якому та твердому небі, задній стінці глотки, мигдаликах, надгортаннику та гортані.
Тактильна сенсорна система, поряд з пропріорецептивною, зорової та вестибулярної сенсорними системами, «постачає» головному мозку інформацію про становище та рух тіла у просторі, про становище його окремих частин. Крім того, вона відіграє важливу роль в орієнтації людини в навколишньому середовищі (особливо сильно шкірний дотик розвивається у сліпих і сліпоглухонімих, дозволяючи тим самим таким людям уникати впливу агента, що ушкоджує). Завдяки тактильній сенсорній системі здійснюється контакт немовляти з матір'ю, виконання різних ігрових, освітніх і трудових операцій, інтимні відносини між чоловіком і жінкою, відчуття комфорту від одягу, що носиться. У принципі такі приклади, що доводять значення тактильної рецепції, можна перераховувати ще й ще. Скажімо лише одне – порушення цього виду чутливості суттєво знижує адаптаційні можливості людини та позбавляє її багатьох радощів життя. p align="justify"> Особливе місце тактильна сенсорна система займає у ссавців, виконуючи життєво важливі функції - тактильне дослідження навколишнього середовища, харчування, звукоутворення і т.д.
Тактильна чутливість різко підвищена на ділянках тіла, які вкриті волоссям. Це пояснюється тим, що волосся грає роль важелів і посилює передачу подразнення в кілька разів, а так як 95% поверхні людського тіла покрито волоссям, на деяких ділянках ледве помітним, то будь-який дотик до поверхні тіла різко посилюється.
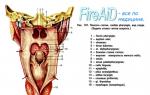
Рецептори відчутного аналізатора.Розрізняють такі основні механорецептори - вільні неінкапсульовані нервові закінчення, вільні нервові закінчення волосяних фолікулів, диски Меркеля, тільця Руффіні, тільця Мейснера та тільця Пачіні.Будова вказаних рецепторів по-різному, вони розподілені нерівномірно і локалізуються на різній глибині шкіри. Перші два типи рецепторів відносяться до первинних (є закінченнями дендриту чутливого нейрона), інші – до вторинних (є інкапсульованими спеціалізованими клітинами, що трансформують механічний вплив на рецепторний потенціал, який передається на дендрит чутливого нейрона). Розглянемо окремі види рецепторів (рис. 4.1).
Мал. 4.1. Схема будови та положення механорецепторів у шкірі, на не покритих волоссям (А) та волосистих (Б) ділянках шкіри.
Вільні неінкапсульовані нервові закінченняє найпоширенішими рецепторами шкіри. Вони знаходяться в основному в сосочковому шарі дерми - зазвичай йдуть вздовж дрібних судин і є розгалуження дендриту аферентних нейронів. Спочатку їх вважали рецепторами болю, але в даний час їх розглядають як полімодальні рецептори, що відповідають і больові, і температурні, і механічні стимули. Це рецептори, що повільно адаптуються: продовжують відповідати весь період часу, поки діє стимул.
Вільні нервові закінчення волосяних фолікулівтакож є розгалуження дендриту аферентного нейрона, які обплітають волосяну сумку. Зазвичай фолікул отримує волокна від кількох чутливих нейронів, але водночас той самий дендрит чутливого нейрона іннервує кілька волосяних фолікулів. Волосся, як було вже зазначено вище, служить важелем, що посилює подразнення нервових закінчень, що пояснює високу чутливість волосся до дотику. Зазначені рецептори реагують переважно на легкий дотик та здійснюють просторове та тимчасове тактильне розрізнення.
Диски (клітинні комплекси) Меркеляє видозміненими епітеліальними клітинами, з якими утворюють синапси дендрити аферентних нейронів. Знаходяться в базальному шарі епідермісу та частково в сосочковому шарі дерми у вигляді невеликих скупчень. Особливо багато дисків Меркеля у ділянках шкіри із високим чутливістю, тобто. у голій шкірі пальців рук і на губах. У покритій волоссям шкірі вони лежать в особливих дзвонових тільцях, що видаються над навколишньою шкірною поверхнею (у кожному такому утворенні, званому тактильним тільцем Пінкуса-Ігго, знаходиться 30-50 клітин Меркеля, пов'язаних з аферентними нервовими волокнами). При висоті близько 0,1 мм і діаметрі 0,2-0,4 мм ці тільця ледь помітні неозброєним оком.
Оскільки диски Меркеля повільно адаптуються рецептори, їх розглядають як пропорційного датчика, тобто. генерація рецепторного потенціалу у яких тим активніше, що сильніше діє подразник. Ці рецептори розглядають як рецептори тиску або сили, оскільки вони сприймають зміну сили механічного впливу.
Тільця (закінчення) Руффінірозташовуються у волосистій частині шкіри – у глибоких шарах епідермісу та у сосочковому шарі дерми. Є веретеноподібною формою капсулу, утворену щільно переплетеними колагеновими волокнами. Усередині такої капсули міститься рідина, в яку занурені розгалуження дендриту чутливого нейрона. Як і диски Меркеля, є рецепторами тиску чи сили.
Тельця Мейснера (тактильне мейснерове тільце)є капсулою конусовидної або овальної форми. Капсула орієнтована перпендикулярно поверхні шкіри. Її стінки утворені численними пластинчастими клітинами, між якими паралельно поверхні шкіри розташовуються терміналі дендриту чутливого нейрона. Ці рецептори знаходяться в сосочках власне коріуму, в ділянках, позбавлених волосся (шкіра долонь, підошв, пальців рук та ніг, а також губи, сосочки молочних залоз та статеві органи). У покритій волоссям шкірі їх менше і тут вони набувають форми рецепторів волосяних фолікулів. Реагують швидкість зміни сили, тобто. є датчиком швидкості.
Тільця Пачіні (пластинчасті тільця, тільця Фатера-Пачіні)- Найбільші і найбільш поширені в організмі рецептори, а тому вони є найбільш вивченими рецепторами шкіри. Розташовуються в гіподермі та частково у глибоких шарах дерми. Крім того, вони виявлені і в сухожиллях та фасціях м'язів, у окістя, у суглобових сумках. Тільце Пачіні нагадує цибулину, складається із зовнішньої капсули, внутрішньої колби та укладеного в неї дендриту аферентного нейрона. Простір між зовнішньою капсулою та внутрішньою колбою, а також усередині колби заповнений рідиною (ліквором). Порушення цих рецепторів відбувається при короткочасному (протягом 0,1 мс) зміщенні капсули на 0,5 мкм.
Тільці Пачіні вважаються датчиком прискорення. Завдяки своїм властивостям вони сприймають незначну деформацію шкіри при зіткненні з різними предметами і грунтом, тобто. з якою швидкістю зміщується шкіра.
Таким чином, більшість тактильних рецепторів локалізовано, як правило, у глибоких шарах епідермісу та сосочковому шарі коріуму. Всіх їх можна розділити на три основні типи:
- рецептори тиску (датчики сили),які поводяться як пропорційні датчики, тобто. генерація рецепторного потенціалу у яких тим активніше, що сильніше діє подразник. Тому ще називають пропорційними рецепторами. Це вільні неінкапсульовані нервові закінчення, диски Меркля, тільця Руффіні;
- рецептори дотику (датчики швидкості)реагують швидкість зміни сили, тобто. швидкість вдавлення стимулу, тому здійснюють просторове та тимчасове тактильне розрізнення. Це тільця Мейсмера та рецептори волосяних фолікулів;
- рецептори вібрації (датчики прискорення)- Тільця Пачіні - реагують на зміну швидкості механічного впливу.
Однак при цьому необхідно підкреслити, що механічні стимули, які зазвичай діють на шкіру, одночасно збуджують у тій чи іншій мірі кілька типів механорецепторів залежно від характеру стимулу. Тому відчуття, що виникають, не можна приписати рецепторам одного типу. Відповідно у повсякденному житті важко визначити різницю між відчуттями тиску і дотику.
Принцип роботи механорецепторів. Незалежно від того, що одні рецептори реагують на зміну сили, другі – на швидкість зміни цієї сили, а треті – на другу похідну зміни сили, у всіх випадках принцип роботи рецептора полягає в тому, що під впливом механічного стимулу в мембрані рецептора змінюється іонна проникність що призводить до появи рецепторного потенціалу. Він викликає виділення медіатора, що супроводжується появою генераторного потенціалу дендритах чутливого нейрона, завдяки якому змінюється частота генерації потенціалів дії. Відмінності у сприйнятті механічних стимулів визначаються швидкістю адаптації тактильних рецепторів. Так, рецептори, що швидко адаптуються, є датчиками швидкості, або прискорення, а повільно адаптуються - датчиками зміни сили. В той же час швидкість адаптації визначається структурою рецепторів – наявність складно організованої капсули рецептора підвищує швидкість його адаптації (укорочує рецепторний потенціал), оскільки така капсула добре проводить швидкі та гасить повільні зміни тиску. Адаптація механорецепторів шкіри має велике значення – завдяки цій властивості рецепторів ми перестаємо відчувати постійний тиск одягу, звикаємо носити на рогівці очей контактні лінзи, а на носі – окуляри тощо.
Проведення відчутної інформації до центрального відділу.Сенсорна інформація від механорецепторів шкіри передається в кору великих півкуль по нервових волокнах. тонкого та клиноподібного пучків, що утворюють лемніскову систему.Тонкий пучок, або пучок Голля, несе інформацію від рецепторів нижніх кінцівок та нижньої частини тулуба, а клиноподібний пучок, або Бурдаха, - від рецепторів верхніх кінцівок та верхньої частини тулуба. Обидва шляхи складаються з аферентних нейронів (вони перебувають у спинномозкових вузлах), аксони яких вступають у задні роги спинного мозку і, не перериваючись, прямують у складі задніх стовпів до тонкого (ядро Голля) та клиноподібного (ядро Бурдаха) ядер довгастого мозку. Аксони нейронів цих ядер повністю перехрещуються лише на рівні олив, утворюючи медіальний лемніск (петлю), і, піднімаючись далі, закінчуються у специфічних ядрах таламуса, які з-за свого анатомічного становища називають вентробазальним комплексом. Нейрони специфічних ядер таламуса посилають свої аксони до соматосенсорних зон кори великих півкуль (SI і SII). Поразка задніх стовпівклінічно проявляється у втраті м'язово-суглобового почуття, вібраційної та тактильної чутливості на стороні ураження при збереженні больової та температурної чутливості.
Лемніскова система проводить точну (за силою і місцем впливу) і складну (про тиск, дотик, вібрацію і рух у суглобах) інформацію з великою швидкістю (до 80 км/с).
Для всіх складових лемніскової системи (задніх стовпів спинного мозку, тонкого та клиноподібного ядер, таламічних ядер та кіркових областей) виявлено чітку топографічну організацію проекції шкіри, тобто. кожна ділянка шкіри контрлатеральної частини тіла займає певну зону, площа якої залежить від функціонального значення цієї частини тіла.
Частина імпульсів від рецепторів тиску шкіри і слизової оболонки передається в кору великих півкуль і по вентрального спиноталамічного тракту,який проходить у складі передніх стовпів спинного мозку. Однак у спиноталамічній системі на рівні таламічних ядер правильна соматотопічна організація представництва поверхні тіла відсутня.
Особливе місце у передачі сенсорної інформації від механорецепторів відводиться V парі черепно-мозкових нервів. трійчастому нерву, який у своїх трьох гілках містить аференти, що йдуть від обличчя та порожнини рота. Він іннервує шкіру, зуби, слизову оболонку порожнини рота, язик і рогівку. На час народження трійчаста система вже добре розвинена і проводить сенсорні сигнали, які запускають харчову поведінку. Саме вона забезпечує перше сенсорне знайомство новонародженого з навколишнім світом.
Корковий рівень аналізу тактильної інформації.Інформація від нейронів специфічних ядер таламуса спочатку надходить у дві проекційні соматосенсорні зони кори великих півкуль (SIіSII). Зокрема, інформація від нейронів вентробазального комплексу направляється (контрлатерально) до першої проекційної зони, яка у приматів та людини знаходиться у постцентральній звивині (SI). Від нейронів задньої групи ядер таламуса інформація переважно надходить (контрлатерально та іпсилатерально) до другої проекційної соматосенсорної зони кори (SII), яка розташована в області сильвієвої (латеральної) борозни (поряд із слуховою зоною). Від цих двох проекційних соматосенсорних зон інформація надходить у передні та задні асоціативні зони кори.
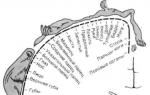
Перша проекційна соматосенсорна область, локалізована в постцентральній звивині (первинні зони - 1-е та 3-е поля по Бродману, вторинні зони - 2-е та 5-е поля), є, по суті, місцем закінчення лемніскового шляху та ядром тактильного аналізатора . Вона відрізняється від інших областей кори дуже високим ступенем топографічної організації (проекції різних областей поверхні тіла на соматосенсорну кору здійснюються за принципом "крапка в крапку"). Таке явище називають соматотопією, або топографічнимпредставництвом. Про наявність соматотопії свідчить той факт, що при подразненні поверхні шкіри короткими точковими дотиками первинна відповідь у соматосенсорній корі локалізується в обмеженій ділянці. Показано, що розміри представництва відповідних ділянок поверхні шкіри у людини та приматів у постцентральній звивині пов'язані не з величиною поверхні їхнього тіла, а залежать від біологічної значущості інформації, що сприймається тією чи іншою ділянкою шкіри. Наприклад, у людини представництво губ, обличчя та кистей рук у цій зоні за площею набагато більше, ніж представництво тулуба та нижніх кінцівок. Це дає специфічний малюнок сенсорного гомункулюса – відображення тіла людини у постцентральній звивині (рис. 4.2). Характер цього малюнка свідчить про високу чутливість і тонке розрізнення для цих частин тіла, а також їх біологічну значущість. Так, під час огляду будь-якого предмета, коли треба з'ясувати форму, наявність шорсткостей тощо., ми погладжуємо цей предмет, тобто. торкаємося його поверхні шкірою нашої долоні.
Коли кора в ділянці SI зруйнована ушкодженням або видалена з терапевтичною метою, виникає дефіцит сприйняття. Стимуляція шкіри може бути сприйнята як така, але здатність локалізувати її і розпізнати просторові деталі подразника порушується. Так, наприклад, хворі не могли навпомацки відрізнити овал від паралелепіпеда (у клініці це порушення носить назву астерогнозис). Ступінь дефіциту залежить від розмірів пошкодженої ділянки кори. Тут знову виступає соматотопічна організація. Однак треба сказати, що після досить довгого проміжку часу подібні порушення стають слабшими. Таке поліпшення, мабуть, обумовлено здатністю інших областей кори (наприклад, 5-го поля тім'яної кори, сусіднього з SI) прийняти він функції SI.

Рис.4.2. Схема чутливого гомункулюса (соматотопічна організація соматосенсорної кори SI людини).
Розріз півкуль (на рівні постцентральної звивини) у передній площині. Позначення показують просторове представництво поверхні тіла в корі, встановлене на підставі локальної стимуляції мозку хворих, що не сплять.
Аналіз інформації від тактильних рецепторів у першій соматосенсорній зоні здійснюється нейронами, об'єднаними у вертикальні колонки, які можна як своєрідні функціональні одиниці, чи блоки кори. Кожна така колонка, отримуючи інформацію від рецепторів однієї і тієї ж модальності, що знаходяться на тому самому рецептивному полі шкіри, проводить цей аналіз за участю спеціалізованих нейронів, число яких в колонці досягає 10 5 . Кожен із цих нейронів «налаштований» на певну ознаку, наявність якого в інформації, що надходить, викликає збудження відповідного нейрона. Завдяки діяльності колонок мозок отримує інформацію про всі властивості стимулу, що впливає на відповідну ділянку шкіри.
До другої соматосенсорної зони кори великих півкуль (SII), розташованої в області сильвієвої борозни поблизу слухової зони (40-і та 51-і поля), надходять імпульси від тактильних рецепторів шкіри «своєї» і протилежної сторони. Ця зона містить точне та детальне представництво поверхні тіла, як і перша соматосенсорна зона, з тією відмінністю, що проекції обох половин тіла в другій соматосенсорній зоні повністю перекриваються, завдяки чому відбувається об'єднання та порівняння інформації, що надходить від правої та лівої половини тіла, тобто . має місце білатеральнесоматотопічний представництво. Як вважають, SII спеціально відіграє роль сенсорної і моторної координації активності на двох сторонах тіла (наприклад, хапання або обмацування обома руками). Не виключено, що друга соматосенсорна зона може, крім того, контролювати аферентну передачу сигналів у таламічних ядрах.
Від первинних і вторинних проекційних зон кори інформація від тактильних рецепторів надходить у передні (фронтальні) і задні асоціативні зони кори, завдяки яким завершується сприйняття, тобто. відбувається упізнання образу (акцепція сигналу). Це реалізується за участю спеціальних нейронів («бабусиних» нейронів), які проходять «навчання» у процесі індивідуального розвитку людини.
В цілому роль соматосенсорної зони кори полягає в інтегральній оцінці соматосенсорних сигналів, у включенні їх у сферу свідомості, полісенсорний синтез та сенсорне забезпечення вироблення нових рухових навичок. Видалення чи пошкодження соматосенсорних зон кори призводить до порушення здатності локалізувати тактильні відчуття, які електростимуляція викликає відчуття тиску, дотику, вібрації і сверблячки.